 |
|
|
|
|
|
In 2016 two atomic structures of mammalian Complex I at 3.9A (Fiedorczuk et al, Nature) and 4.2A (Zhu et al, Nature) were released by two groups. The update is coming!
StructurePrimary structure: Bovine heart complex I is composed of at least 45 different subunits whose primary structures have been determined. Of these, 7 are encoded by mitochondrial DNA (mtDNA) and the remaining 36 are encoded by nuclear DNA. Recently, entire DNA sequences of human, C. elegans and rice have been determined. The primary structures of the complex I subunits in these eukaryotes are also available. In addition, the primary structures of most of complex I subunits in Neurospora crassa and Yarrowia lipolytica mitochondria are also available.
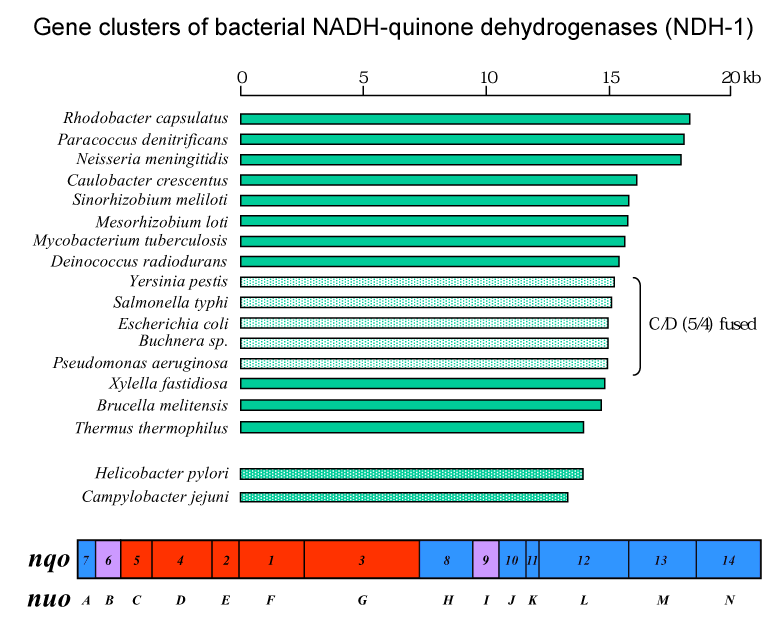
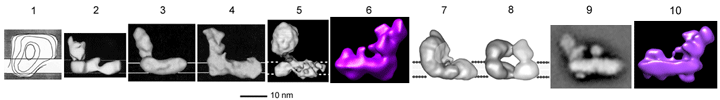
Subunit composition of complex I/NDH-1 from several organisms is compared in this TABLE. Mitochondrial complex I houses at least 29 additional subunits compared to bacterial NDH-1. These subunits have been referred to as accessory (or supernumerary) subunits. Initially, these subunits were considered to be not essential for structure and function of mitochondrial complex I. However, there are several reports on possible functions of the accessory subunits that suggest some of these subunits are indeed essential (see Hot topic 3 below). 3D structure: Complex I consists of 2 major segments. One is the peripheral segment which protrudes into the mitochondrial matrix (or bacterial cytosol) . The other is the membrane segment. The peripheral segment is a "catalytic" domain and is composed of 7 (bacteria) to 12 (mitochondria) subunits. The membrane segment consists mainly of hydrophobic subunits including, in the case of mitochondrial enzyme, all mitochondrial DNA-encoded subunits. Unlike other enzyme complexes involved in oxidative phosphorylation, only available information on the 3D structures of complex I is from the low resolution EM analyses. Click on the image below for detailed description.
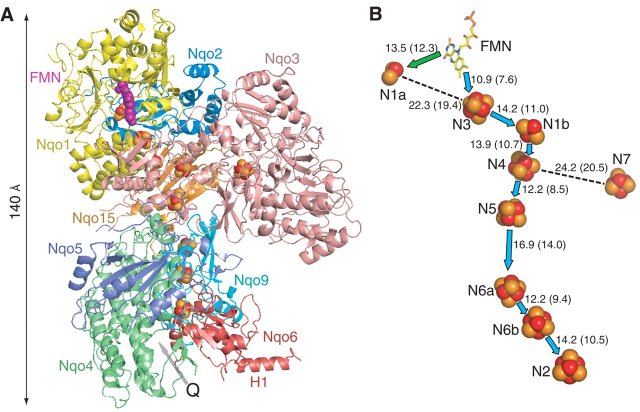
Hot news on the 3D structure: Sazanov and Hinchliffe reported the crystal structure of the hydrophilic domain of complex I from Thermus thermophilus at 3.3 angstrom resolution.
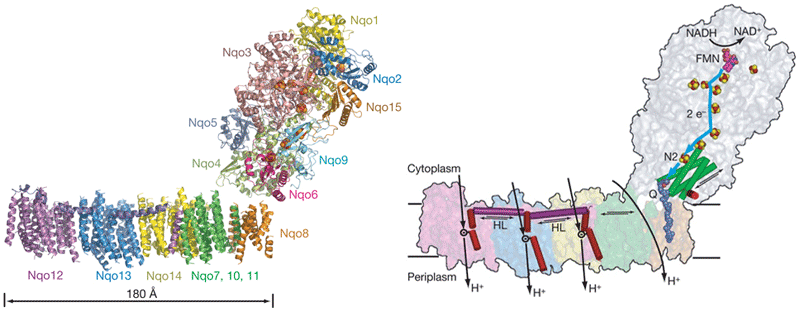
Sazanov's group recently reported the structure of the entire complex I from Thermus thermophilus at 4.5 angstrom resolution. ( Efremov, R. G., Baradaran, R. and Sazanov, L. A.) *
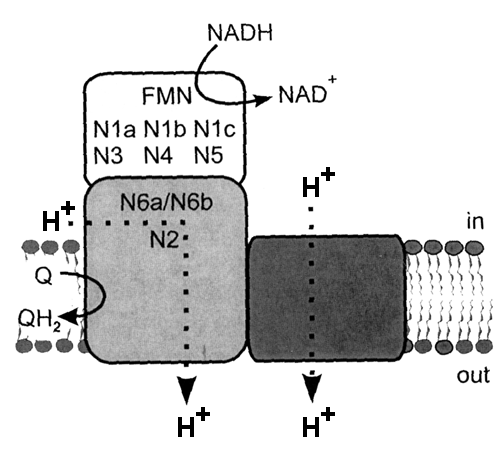
Location of cofactors: Mitochondrial complex I and bacterial NDH-1 contain one noncovalently bound FMN and 8-10 iron-sulfur clusters. On the basis of terminology by Ohnishi's group, N1a and N1b are binuclear clusters and N2, N3, N4, N5, and N6 are tetranuclear clusters. The NDH-1 in certain bacteria contain an additional tetranuclear cluster (designated N7) and N6 containing two tetranuclear clusters. Clusters N1a and N1b are located in subunits Nqo2/NuoE/ FP24k and Nqo3/NuoG/IP75k, respectively. Center N3 is located in subunit Nqo1/NuoF/FP51k. Clusters N4, N5, and N7 are present in subunits Nqo3/NuoG/IP75k. The cluster N6 (2[4Fe-4S]) is coordinated by subunit Nqo9/NuoI/TYKY. Center N2 is probably housed in subunit Nqo6/NuoB/PSST. The details are shown in this TABLE. MechanismVarious investigators proposed their own energy coupling mechanism of complex I. These hypotheses can be divided into at least two groups. One is direct ion coupling mechanism to the electron transfer. The other is indirect coupling mechanism.The direct coupling mechanism can be further divided into at least two groups. One is that quinone(s) is simply involved in proton-translocation (Degli Esposti, Vinogradov, and Stueber). The other is akin to the Q-cycle mechanism in complex III (Brandt and Dutton). For details please read the references by the investigators.
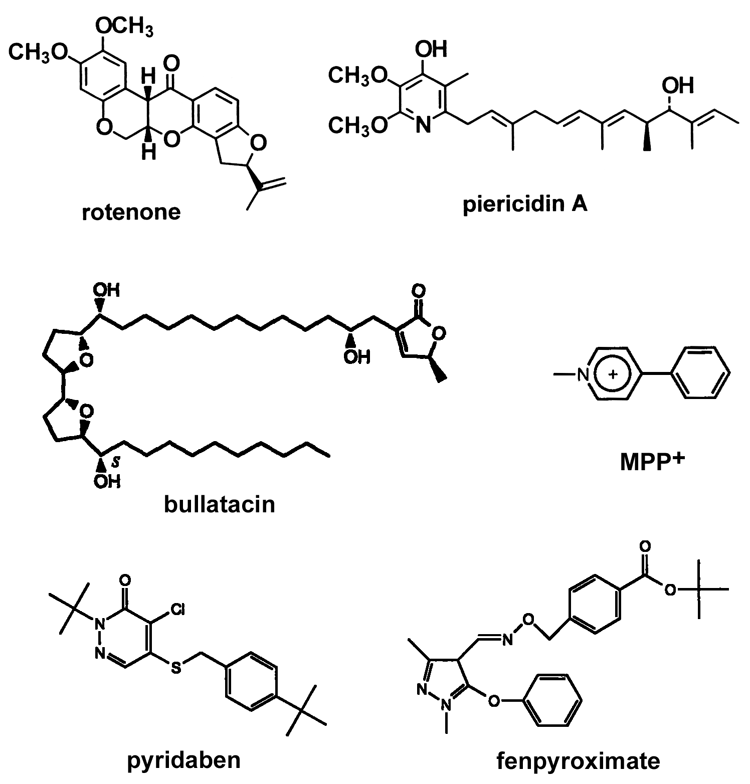
InhibitorsThe most well-known inhibitor specific for complex I is rotenone. However, there are a number of compounds, both naturally-occurring and synthetic, that are potent inhibitors of complex I. Some typical inhibitors are shown in this TABLE and their structures are in this FIGURE.Complex I-related diseasesAs described in the Overview section, there are a number of diseases caused by defects of Complex I. Although it is not covered extensively here, there are excellentweb sites you can refer to.(OLD!) Hot topics and unsolved questionsTopic 1: Where are proton-translocating site(s) and quinone-binding site(s)?
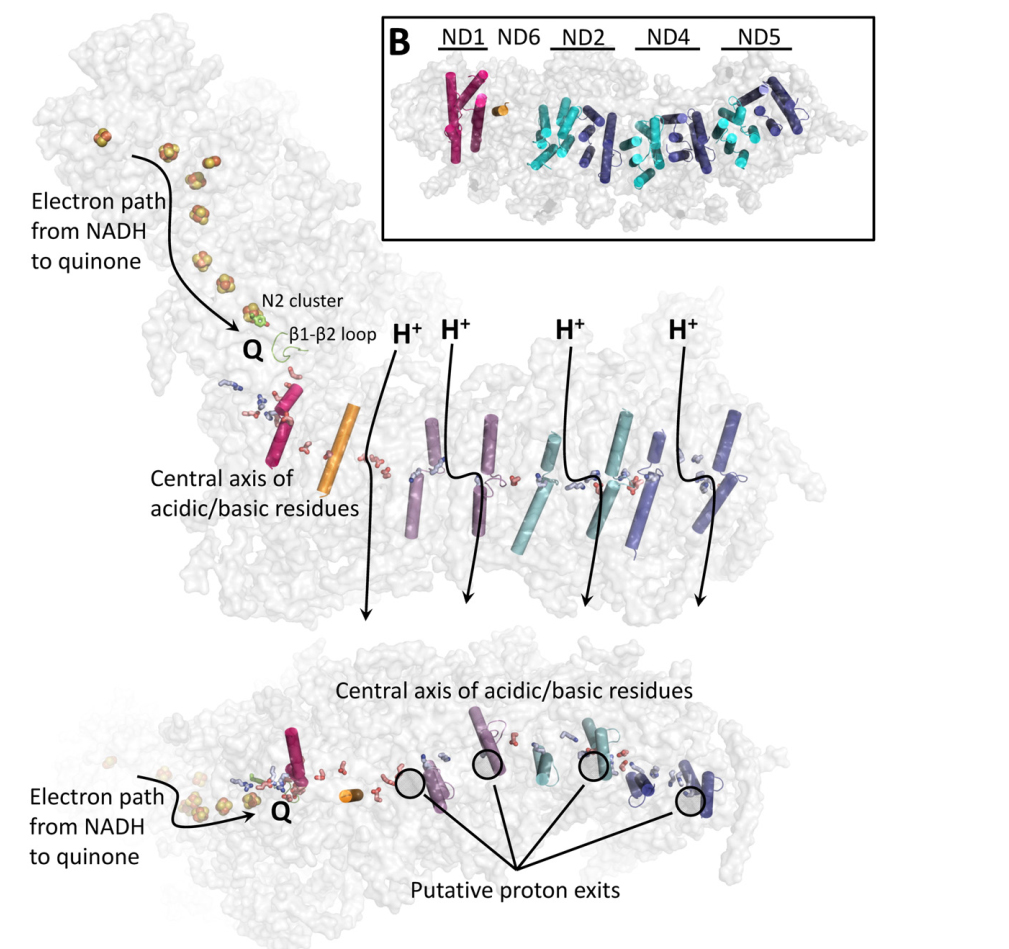
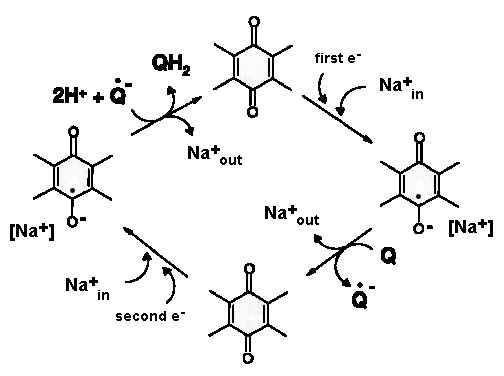
UPDATED: Look at the figure (from PMID:26921811). Quinone binding site is formed by the 49-kDa and PSST subunits. Ubiquinone accepts electrons from the FeS cluster N2 which is located ~30 Å above the membrane plane. Conformational changes linked to the redox chemistry of ubiquinone are proposed to induce proton translocation at four sites of the membrane arm (subunits ND1/ND6, ND2, ND4 and ND5). Topic 2: Can Complex I pump not only protons but also sodium ions? OLD: Recently, Steuber's group demonstrated that complex I in certain bacteria can work as a sodium pump. A question arises whether this feature is common to complex I of other sources. UPDATED: No direct proofs were found to support this claim. The published data showing thta complex I pumps Na+ in K. pneumoniae by Steuber (PMID:10417649, PMID:12110677, PMID:12538874, PMID:11695827) can be partially explained by contamination of the preparation by Na+-translocated NADH:quinone reductase (Bertsova&Bogachev 2014). No indication for sodium translocation by mitochondrial complex I was found for the yeast enzyme (PMID:17094937, PMID:16289468). However, there is a publication that teh D-form of bovine complex I can posess s Na+/H+ antiporter activity (PMID:22854968) Topic 3: Are accessory subunits in mitochondrial complex I really accessory? OLD: Weiss'group and Walker's group reported that acyl carrier protein of the bacterial fatty acid synthesis system is a subunit of complex I. Hatefi's group and Schulte's group showed that the HP39k subunit binds NADPH. Papa's group reported that the IP18k subunit is phosphorylated and this phosphorylation is involved in regulation of complex I activity. Videira's group suggested involvement of some subunits in the assembly of N. crassa complex I. Recently, Scheffler's group demonstrated that the MWFE subunit is essential for complex I activity. These results raise a question as to whether "the accessory subunits" are really accessory. UPDATED: There is a significant progress in identification of structure and possible function of so-called accessory subunits of complex I. Please look here: PMID: 27626371, PMID:24059519 , PMID:21711245, PMID:21115482. |
|
{kind=link}
{kind=link}
{kind=link}